





作者:古明地恋
本篇是关于白垩纪大灭绝,你想知道又找不到的内容。
小行星以哪个方向,何等速度,怎样能量撞上地球?小行星是什么质地?陨石坑有多大?炸毁四分之一地球的冲击波?波及全球的大火?57级大的狂风?11级地震和300米高的海啸?撞击冬天持续了多久?恐龙,沧龙,翼龙,菊石在撞击后生活了多久?相信这篇文章会解决你对白垩纪末大灭绝的许多好奇。

序:撞击理论
20世纪70年代末,加州大学伯克利分校的一个科学团队,一个戴着眼镜的核物理诺贝尔奖获得者:路易斯。沃尔特。阿尔瓦雷斯,提出了一个理论。在《地球的沉积物》一书中,他提出在6600万年前的白垩纪﹣古近纪灭绝事件(k-pg)界线存在异常高的铱含量,而这种重金属很少出现于地球表面,却在陨石中十分普遍。由此,一颗小行星造成恐龙的灭绝的主张被提出。
科学家们起初对此表示怀疑。在此之前,假说通常认为火山或冰川是造成这场大灭绝的主要原因。然而,在100多个出现白垩纪灭绝遗迹的地方都发现了大量的铱,支持了阿尔瓦雷斯的观点。那么,这个理论被承认的最后一步,就是找到这个陨石坑了。
为此,阿尔瓦雷斯团队在地球上孜孜不倦地寻找一个符合他们要求的谷地。1990年6月,就在阿尔瓦雷斯发表声明的20年后,地质学家们在墨西哥希克苏鲁伯镇附近的尤卡坦半岛北角发现了一个巨大的陨石坑,这个陨石坑因此得名为“希克苏鲁伯陨石坑”。
通过对这个陨石坑和其他遗迹的分析,我们得以还原66.043百万年前浩劫发生的那一刻,以及紧随其后数十年的情景。
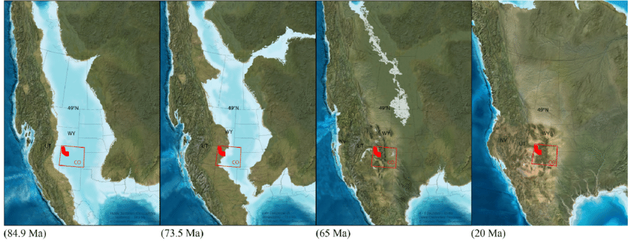 从8490万年前到2000万年前北美洲的西部内陆海道由宽阔到消失的过程。
从8490万年前到2000万年前北美洲的西部内陆海道由宽阔到消失的过程。Stage1:撞击前夕
白垩纪末期全球持续变冷,海平面下降。晚白垩世海拔高峰值为50-70米,随之而来的是坎帕——马斯特里赫特阶长期的跌落。在早白垩纪—中白垩纪,一片浅海:著名的“西部内陆海道”覆盖着北美洲的中西部地区,但因为持续板块运动形成落基山,导致大陆的拉伸和断裂,覆盖的海水撤退。到了灭绝前夕,内陆海道仅剩一狭窄延伸的小片区域。而到了古近纪,仅余其中被称为炮弹海(Cannonball Sea)的一小片残余。
 白垩纪的北美洲及恐龙,图2为撞击前夕的地图
白垩纪的北美洲及恐龙,图2为撞击前夕的地图白垩纪末期,印度西部下方发生了大规模火山喷发,火山喷发主要分为三个阶段:第一次发生在C30n(地层名),第二次发生在C29r(地层名),第三次发生在C29n(地层名)。第二阶段可能开始于在白垩纪﹣古近纪灭绝事件(K-Pg)边界之前约40万年,为最大的一次并形成了高达80%体积的德干地盾。德干火山相关的全球气候变化,很可能与白垩纪-古近纪界线标志性的大灭绝有关——无论是好事还是坏事。

无论如何,德干火山作用可能在白垩纪﹣古近纪灭绝事件(K/Pg)大灭绝事件之前200 kyr(1kyr=1000年)造成了地表海洋酸化,尤其影响了带有钙质外壳的生物——有孔虫,颗石藻和菊石。岩石中的化石记录证明,在希克苏鲁伯陨石撞击之前,海洋无脊椎动物的物种数就已经在逐渐减少。在灭绝前夕,平旋菊石已经从浅海区域消失,仅存少量快速游动类型分布于远洋海域。
也有文章表明,德干火山带来的热量反而从撞击后的长期冬天救回了一些生命。

Stage2:撞击瞬间(撞击10分钟内)
66.043百万年前,小行星以60度的水平倾角,自东北方向撞向地球。
这次撞击留下了直径近300公里的撞击坑,表明其可能是自40亿年前早期撞击结束以来,在太阳系内部产生的最大的撞击结构之一。
结果表明,希克苏鲁伯陨石坑瞬时直径(D)的合理估算值约为170士25 km。由此计算和实验得出的陨石坑尺度关系表明,撞击事件的深度在17~20公里之间,瞬时陨石坑的深度在45~60公里之间。直径约300公里的希克苏鲁伯多环盆地记录了近40亿年前后期重轰炸期以来太阳内部最大的碰撞之一。在经过充分研究的内行星和卫星表面上,唯一被发现的大小与之相当的盆地是金星上直径280公里的米德盆地。自从大约10亿年前多细胞生命的发展以来,地球可能还没有经历过如此规模的另一次冲击。
这颗小行星由碳酸盐(方解石)和蒸发岩(硬石膏)组成,有7.5英里(约12km)宽,一颗小行星的撞击所需直径约为12公里,时速约为普通速度20公里/秒;彗星所需的直径范围在10-14公里,撞击速度要提升到50公里/秒。这个速度大概相当于子弹速度的20倍~50倍,释放的能量比目前测试过的最大核弹爆炸能量的100万倍还要大。
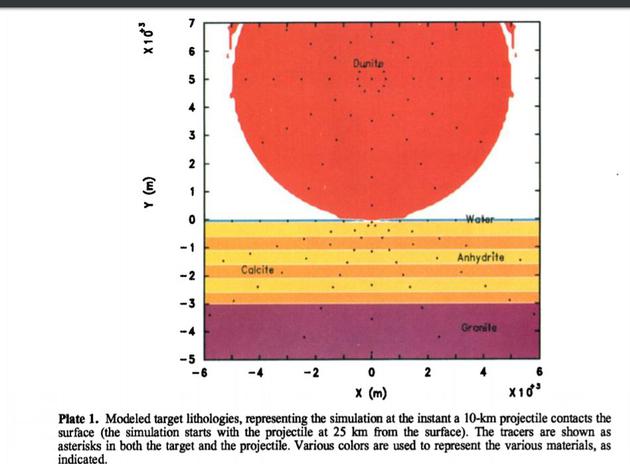 小行星的大小,在10~25千米不等,若是彗星则50km
小行星的大小,在10~25千米不等,若是彗星则50km小行星着陆的位置——希克苏鲁伯,在撞击当天是覆盖着数米深海水的浅海。在撞击的刹那,地表物质便被冲击波掀起,产生巨大的羽流。热量使海水瞬时蒸发,膨胀的羽流便最初主要由浅海被蒸发的水蒸气构成。羽流直冲云霄,巨大的混合在内部进行,在撞击地上方形成一个烟囱状的,含有各种来源于浅海,沉积层,生物体,以及后来陨石破裂的弹射体的烟柱。物质以每秒几公里的速度从羽流的下部被抛到上部,稍后,烟柱开始水平膨胀。
在撞击后大约30秒后,陨石坑大约达到了三分之一的瞬态大小,而羽流已经超出了平流层(离地表10km至50km),继续扩展,水平上达到了撞击坑大小的3倍。
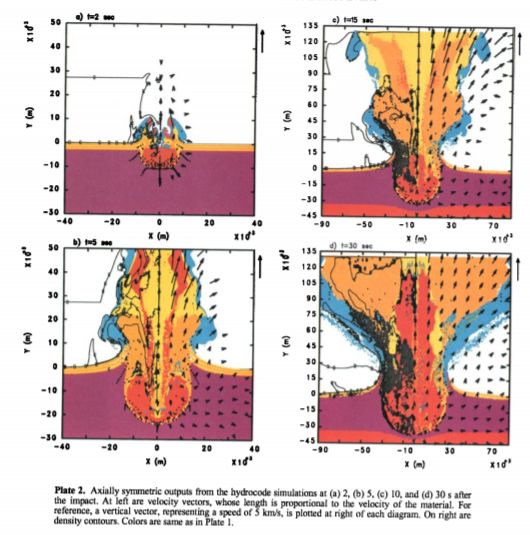 小行星撞击的步骤及产生的羽流
小行星撞击的步骤及产生的羽流小行星只有微不足道的弹丸部分在撞击口外膨胀。撞击表面后,小行星并没有立即碎裂,而是砸碎地壳,钻入上地幔,在瞬间在地底形成了大约100公里直径的瞬时空腔。小行星钻入地底深度深达30-40千米(有文章表示可达70km深)。
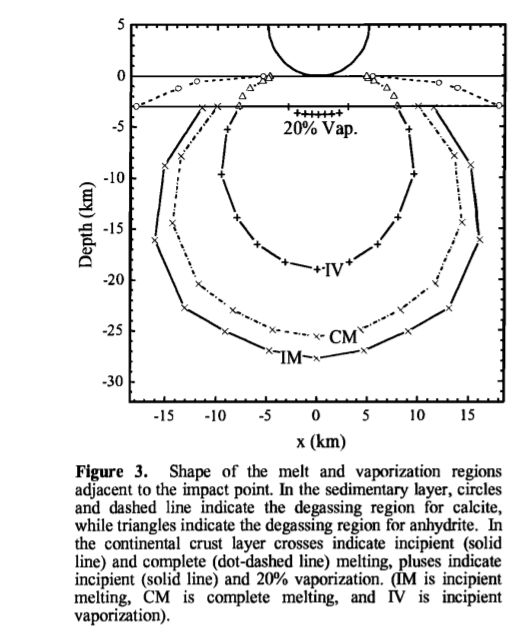 小行星钻入地底的熔岩和空腔范围
小行星钻入地底的熔岩和空腔范围紧接着,巨大的海啸席卷了墨西哥湾,撞击附近的海岸线,并且辐射,穿越原加勒比海和大西洋盆地。海啸在墨西哥湾沿岸发生时高达100到300米,横冲直撞冲进300公里外的内陆,并将海底沉积物冲到海底深处500米。
 被冲击翻到k-pg界线的中白垩纪有孔虫
被冲击翻到k-pg界线的中白垩纪有孔虫撞击事件同时产生了辐射整个北美洲的爆炸冲击波,在冲击地附近引发超过每小时1000公里的56级大狂风(地球上最大的风级17级,为200-220km/h,风速高时20km/h为一级),冲刷土壤,撕碎植物和任何生活在附近的生物。直径约为3000公里的地区被空中爆炸破坏。(PS:地球半径 6371.012Km,爆炸波及长度相当于地球的四分之一)。
从火山口升起的烟羽温度超过10000摄氏度,相当于太阳表面温度的2倍,在距离1500 - 4000公里以内引发巨大火灾。如此高的温度对生活在这个范围内的动物来说是毁灭性的。热脉冲相对较短,只持续了5-10分钟,所以有些生物如果受到庇护,就能逃脱这种特殊的影响。当撞击喷出物落下时,产生的熔岩火雨持续了3 - 4天。
 小行星撞击的火焰温度相当于太阳表面温度的2倍
小行星撞击的火焰温度相当于太阳表面温度的2倍撞击还生产了破坏臭氧的氯和溴。超过5个数量级的,混合了溴和其他反应物的氯注入到同温层,比破坏今天的臭氧层所需要的还要多。大气中产生的氮化物也具有破坏臭氧的能力。对臭氧层的影响可能持续了好几年。
Stage3:火焰地球
陆地灭绝模式(数小时——数天)
第一个小时后,撞击引发的>11级的地震和可达300米高的巨大海啸是主要的作用,也即所谓的“火球阶段”。这次撞击引爆了数万吨的岩石,还令小行星的残留物被冲击到了大气层,其中某些元素进入了轨道,而其余部分变成密集燃烧着的流星返回地面,包括由更大的喷出物回流而产生的强烈的热脉冲,造成全球森林大火。红外光谱表明,喷出物分布在全球范围内可能导致了短期大面积的大气变暖,地面温度上升引起树木自燃,在撞击后的数周内烧掉了地球上半数以上的植被。

火灾可能从北美南部生成,但白垩纪﹣古近纪灭绝事件(k-pg)界线上的全球岩屑层足以表明整个陆地生物圈都被烧毁了。初步模拟表明,喷出物重返大气层会引起全球红外(IR)脉冲足以在希克苏鲁伯撞击几小时内引发全球大火。大火假设解释了北美古新世早期的陆地生存模式,因为所有幸存的物种都必须在地下或水中躲避高温和火焰。一些地区比较幸运,在撞击发生时,现场被积水覆盖,成为美洲陆地上少数未被烧焦的地区,留下了缺乏木炭和存在未烧焦的有机物的遗址。
 来自撞击地层的烟灰遗迹
来自撞击地层的烟灰遗迹全球野火将大部分陆地生态系统化为灰烬,从而导致动物大量死亡。在短期内,北美的昆虫似乎消失了,因为北达科他州化石记录中的树叶被昆虫破坏的频率急剧下降。目前还不清楚这些昆虫是否直接死于撞击事件的高温火焰,或是因为它们的寄主植物被杀死而死亡。有穴居特性的类群,如哺乳动物,可以为它们提供免遭最初数分钟内高温的庇护,从而免于灭绝。
 生活在树上的昆虫自然难免一死,而洞中的生物可以免于高温冲击的劫难。
生活在树上的昆虫自然难免一死,而洞中的生物可以免于高温冲击的劫难。随着植物和地表有机碳的燃烧,大量的二氧化碳,CH4和H2O被排放到空气中,其中一些气体是直接来自于小行星(碳酸盐),而其余来自地球本身。撞击产生了总体积约8万立方千米的喷出物,包括从350亿吨到3500亿吨二氧化碳,40亿吨到560亿吨的硫,以及200亿吨到1400亿吨的水蒸气。
撞击事件改变了全球的环境,即使不是持续1000年,至少也会持续几年。
硫化合物可能是最多的注入平流层的重要气候活跃气体。这次撞击释放的硫的数量比任何已知的火山喷发都要高几个数量级,而喷发的水足以对地球气候产生突然而重大的扰动。
喷出物在撞击之后造成了高层大气的强烈升温,而这种作用可能只持续了一小段时间。几个小时后,硫酸盐连续形成,并保持于平流层中,与水蒸气反应生成产生稳定,长寿命的硫酸盐气溶胶。气溶胶通过吸收长波辐射打乱了平流层的热状态,冷却地球表面,显著扰乱全球多年来的气候,大气环流因此中断了好几年。
酸雨是撞击事件的二次产物。在撞击之后,这场雨可能下了几天,几个月到几年。首先是大气受到撞击事件的冲击加热,产生硝酸雨。这次撞击事件产生了约为1×10^15摩尔的硝酸酸雨,另一个约为3×10^15 mol的硝酸可能是由冲击产生的森林大火产生,而硫酸则是由小行星本身及其后的熔岩,森林火灾引起。
 酸雨会进一步损害植物,污染水中生物,导致进一步的灭绝。
酸雨会进一步损害植物,污染水中生物,导致进一步的灭绝。硫酸雨和硝酸雨的结合也不足以使海洋盆地酸化,但它在浅水或缓冲差的河口和大陆上的影响是复杂的。即使所有的硫在撞击中被释放出来,海洋表面的海水酸化也似乎不太可能引发海洋生物的大规模灭绝,但另一方面,在陆地和附近浅水环境的生物如果土壤不肥沃,无力缓冲地下水的酸化,酸雨的后果可能很严重。
Stage4:寒冷地球
水域及陆地灭绝模式(几天—两年)
在最初几天的高温和燃烧后,火焰趋于平静,但更大的危机即将到来。
撞击产生的气溶胶和撞击后产生的烟灰和硫酸盐反射高层大气中的太阳光,从而直接给地球降温;以及森林大火燃烧有机质化石的有机烟雾吸收短波辐射,阻止阳光到达地表,导致地表温度下降,暂时抑制光合作用,引起陆地和海洋食物网的全球崩溃。
阳光减少到20%,大概就是这个感觉。PS,矛形小箭石是“中生代最后的游泳动物群落”的主要(唯一)游泳成员。
模型模拟表明,阳光到达地球表面的数量可能仅剩约20%,这意味着能源供应将减少约300 W·m ^– 2,导致全球范围短期内表面温度的严重下降。在短时间内,地球温度下降了几度到几十度。由此产生的温差可能在相对温暖的海洋和寒冷的大气之间引发大风暴和飓风,增加了尘埃飘散在大气中的时间。
 海陆温度分布不均会引起风,而风又增加了尘埃的滞留时间。
海陆温度分布不均会引起风,而风又增加了尘埃的滞留时间。几个月到几十年后,大气稳定下来,灰尘开始像雨点一样落下,并在沉积环境中积累起来。这包括全球公认的小行星衍生的微量元素铂族元素(PGE),如铱的峰。比起粉尘,硫酸盐对环境的影响更大。气溶胶的作用会导致数十年的较低海温,即使在大部分尘埃已经从大气中清除之后。
 小行星撞击的特征铱元素要在几个月到几十年后才沉降下来。
小行星撞击的特征铱元素要在几个月到几十年后才沉降下来。水生环境被水保护,得以免受热量和火的影响,但海洋环境和淡水环境仍然显示出巨大规模的灭绝。
在几个月到几年的一段时间,可能是最少6个月的黑暗期内,撞击冬季便导致了全球范围内浮游植物的大规模死亡。因为水生生态系统与陆地环境不同,强烈依赖于日常光合作用的输出自养生物,浮游植物的损失很可能造成水生生态系统灾难性的死亡和灭绝。水生生态系统中灭绝的其他潜在原因包括环境温度和缺氧由于缺乏光合作用的氧气。
在海洋中,受到酸雨和光照的影响,具有钙质外壳的成岩生物颗石藻和浮游有孔虫在碰撞边界崩溃了。直到古新世最初的丹尼阶,它们的丰度仍然很低。流向海底的有机碎屑也急剧减少,在撞击后大约3myr(1 myr=1百万年)仍没有恢复。由白垩纪﹣古近纪灭绝事件(K-Pg)边界的浮游生物支撑的海洋物种灭绝率最高。
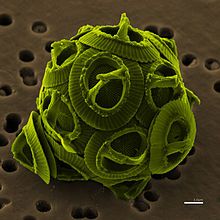 具有钙质外壳的颗石藻,白垩纪十分繁盛,是白垩的主要来源。
具有钙质外壳的颗石藻,白垩纪十分繁盛,是白垩的主要来源。菊石,古生代和中生代最多样化和最丰富的浮游生物捕食者连同以它们为食的巨型沧龙,蛇颈龙和上龙灭绝。随后的古近纪,鱼类向它们曾经占据的生态位扩张。
软骨鱼类失去了大约20%的科,而硬骨鱼失去了大约10%的科。两栖动物几乎没有科层面上的灭绝。所有六个白垩纪晚期海龟科幸存下来。

光合作用的缓慢恢复意味着饥饿是导致海洋物种灭绝的主要原因。海洋物种的灭绝率在远洋物种中更高,因为远洋生物几乎完全依赖浮游植物。而底栖生物有更强的抵抗饥饿的能力,而海底的死亡生物量又被消耗在以尸体为基础的食物链。光共生(zooxanthellate)珊瑚受到大灭绝的影响比偶氮黄藻珊瑚更高。
多细胞变温动物(“冷血动物”)的耐饥能力是体型大小的一个函数(图1:Peters [1983,p 。 42];(Hemmingsen[1960]收集的原始数据)。小型无脊椎动物(10磅至10毫克),包括浮游动物而有些底栖无脊椎动物,生存时间为8—20天。大型无脊椎动物(100克到1公斤)等大型动物,如甲壳纲动物,大型贻贝和许多其他大型底栖动物的生存时间超出了估计的恢复区间的下界6个月;而最大的变温动物,包括特别大的无脊椎头足类,大型鱼类和水生爬行动物 在没有食物的情况下可以生存1到3年。然而,快速自由游动的上龙和沧龙可能是具有较高代谢率的恒温动物,更容易饿死。
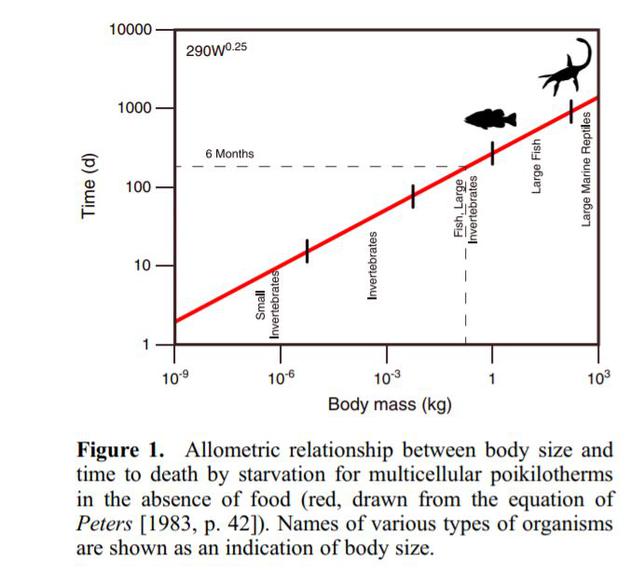 变温动物耐饥饿能力和体型的关系,当然,沧龙和上龙不算。
变温动物耐饥饿能力和体型的关系,当然,沧龙和上龙不算。 中生代的巨大海生蜥蜴沧龙可能也是恒温动物,导致在灭绝中难以生存。
中生代的巨大海生蜥蜴沧龙可能也是恒温动物,导致在灭绝中难以生存。因为耐饥性高,灭绝反而促进了某些海洋生物群体的生存,例如腕足类。在中生代海洋动物群中,腕足类成为了一个次要的组成部分,它们撤退到冷水区,远离温暖和食物丰富的近海。对寒冷的适应可能提高了它们在撞击冬季的生存能力。腕足类以浮游植物,细菌、有机碎屑和有机分子为食。它们可能已经适应了食物匮乏环境中的生活,因此数量在灭绝前后于新泽西州和丹麦相对增加。
 生活在深海中的小嘴贝化石
生活在深海中的小嘴贝化石在海洋里,阳光消失了至少6个月,光合作用可能被抑制了2年之久,但浮游植物此后生产迅速恢复。然而,浮游动物会在撞击后4个月内死于饥饿,因此,完整的食物网并没有在光照重新回归时立即再生。浮游植物在光照恢复后立刻恢复,但被饿死的浮游动物的缓慢恢复可能还有更长的路要走,延缓了生态系统中所有其他动物的恢复。
体型巨大的马斯特里赫特浮游鱼类(>长5米,>重400公斤)可能以浮游动物和磷虾为食,而不是浮游植物。它们庞大的体型可以让它们在两年的撞击冬季存活下来,活到阳光回来,浮游植物在表层水体中反弹的时候。然而,他们并不能等到足够的以浮游植物为食的浮游动物和磷虾的恢复。
 起源于侏罗纪的磷虾类Euphausiacea在中生代就是海洋中的重要食物。
起源于侏罗纪的磷虾类Euphausiacea在中生代就是海洋中的重要食物。这类滤食动物中,菊石是最主要的受害者。大型的aptychoporan ammonites(一种菊石名)生活在马斯特里赫特晚期(Olivero and Zinsmeister)体腔约1.1米长,估计大小这只活的动物重约6公斤,拥有2年左右的饥饿生存极限,但菊石具有浮游营养性的幼虫,需要以浮游植物和浮游动物,因此最终受到浮游动物损失的影响而灭绝。相反,蛸类和鹦鹉螺的幼体是非浮游营养性的,它携带一个卵黄袋,为幼虫提供一个自给营养来源。同样,一些心海胆等(spatangoid)棘皮动物进化出非浮游性的幼虫,因此两者都越过了灭绝。
淡水环境也与陆地同样受到了高温的影响,但水庇护了它们。只有顶端几厘米左右的水体会受到热量的影响,而微小的影响将在几周内完全消散。
撞击产生的能量爆发和随后的火灾可能会烧掉所有暴露的碳,但库存的地下有机碳可以以微粒或溶解的形式运输到淡水生态系统。当然,内陆水域的死亡率同样很高,但灭绝的比例却低于海洋环境,可能是由于淡水类群具有更好的休眠能力,以及河水通过快速流动来抵消氧气需求的曝气效率更高。在温度适中的条件下,地下水提供了丰富的热容,抵消了一部分高温和降温的影响。
几乎所有的淡水科都只有少量的灭绝,与陆地或海洋相反。白垩纪﹣古近纪灭绝事件(K-Pg)界线淡水鲨鱼科没有灭绝,随着离岸距离的增加灭绝率增加,直至45%在开阔的海洋。
马斯特里赫特的10个鳄类科中,有5个海洋科灭绝了,四个淡水科幸存下来,没有淡水科灭绝。甚至生活在淡水中的幼体也能在K-Pg边界生存下来,如离龙目 (包括鳄龙champsosaurs)一直延续到中新世。在淡水中,大型爬行动物(鳄类动物,离龙目,海龟)可能大到足以抵御因饥饿而灭绝。较小的脊椎动物,包括不到一公斤的鱼类和两栖动物就必须依靠其他的生存机制,特别是休眠。幸存的海洋浮游生物,例如甲藻,通常具有形成休眠体的能力,而灭绝率高的浮游生物,如有孔虫类和颗石藻通常缺乏休眠阶段。
 生活在淡水中的似鳄鱼动物离龙成功存活到中新世。
生活在淡水中的似鳄鱼动物离龙成功存活到中新世。此外,以碎屑为食的类群无论是在海洋还是淡水环境中,它们的灭绝率都很低,但是,淡水类群中的食碎屑生物死亡率比海水更低,因为它们可以从更新的土壤中获取更多有机物质。
Stage5:复苏阶段
(几十年—几千年)
气溶胶和尘埃会在几个月到几十年间缓慢沉降,阳光也在6个月——两年后重返地球,达到可维持光合作用的水平。随后,温度开始回升。
之前撞击喷射的温室气体,如二氧化碳,甲烷和其他有机分子帮助了地球的回暖。二氧化碳等气体比那些尘埃和硫酸盐气溶胶的停留时间更长,并在气溶胶和煤烟沉降到地面,冷却结束后引发了温室效应。变暖可能是在一段时间的冷却之后发生的。对加热程度的估计各不相同,根据二氧化碳数据的估算,温室效应上升的温度在1到1.5摄氏度之间。
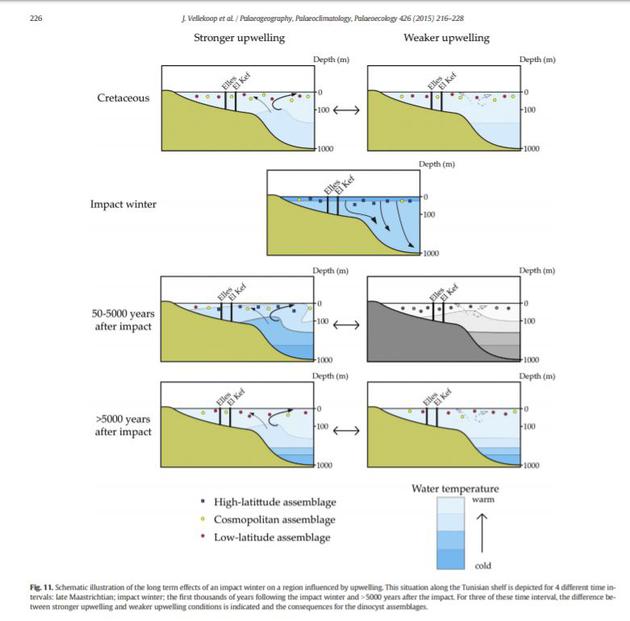 在撞击发生后,海洋表层成了温度最低的区域,深层温度高,恰似冬天的水域;撞击发生50年后,表层温度开始回复正常。
在撞击发生后,海洋表层成了温度最低的区域,深层温度高,恰似冬天的水域;撞击发生50年后,表层温度开始回复正常。随着阳光回归,海洋和淡水的浮游植物迅速恢复,而浮游动物的复原需要更久。然而,在陆地上,植物恢复十分缓慢,因为那里有根植物的生长发育周期至少为一年。在发生火灾的地区,大部分植物都被清除了。
蕨类是北方一些地区,如美国,日本和新西兰的先锋物种。在没有蕨类植物的地区,藻类和苔藓是另外的先驱植被类型。在北美北部,初期的植被由几种类型的蕨类,以及开花的被子植物组成,产生草地。由于传粉动物已经被消灭,那些具有风媒传粉能力而不依赖昆虫的植物从灭绝中活下来的概率更高。落叶树似乎幸存了下来,优于北美的常绿树木,可能是因为他们它们的休眠能力。最终,森林的树冠又回来了。在稍后,由于富含蛋白质的豆类的发展,动物群的重量恢复到较高水平。在130±50 ka(1 ka=1000年)后,美国的碳循环恢复。
 蕨类植物是灭绝后陆地生态恢复的先驱。在此之后,被子植物取代了它们成为优势草本。
蕨类植物是灭绝后陆地生态恢复的先驱。在此之后,被子植物取代了它们成为优势草本。在海洋,随着光照恢复和捕食者,竞争者的消失,个别地方类群开始入侵其他地区。如一个撞击前的近岸有孔虫类群既保持原来的环境生态位,也殖民了远洋环境。复苏在所有的地理位置都不一样。例如,一些软体动物的扩张在撞击地区附近要比在世界的其他地方快得多。沉积喂养的双壳类活了下来,而其他双壳类存活(食肉动物和悬浮捕食动物)的比例为38% - 58%。因为没有了菊石,在古新世,双壳类和棘皮动物经常形成大片的群落。但在新西兰,这一切从开始就是欣欣向荣。新西兰的小型海洋生物化石记录证明,在白垩纪末期该地区没有发生过大规模物种灭绝。事实上,这一地区的生物种类还有所增加,可能是因为导致地球上其他地区动物灭绝的寒流在这里发生了有益物种生存的变化。
Stage6:穿越者们
在非鸟类恐龙灭绝之前,几乎没有证据表明它们的多样性在在白垩纪末期全球范围内长期下降。在地狱河地区的南部恐龙存活了下来,保持多样性和丰富性,没有任何衰落的迹象。在西班牙的特雷普盆地,恐龙幸存了下来,在整个马斯特里赫特阶普遍而多样,在K-Pg边界之前没有明显的减少或局部的灭绝。当然,在北美的白垩纪恐龙群中发现了大型食草动物的多样性减少,也许会使群落更容易受到级联效应的影响。结合撞击事件,我们可以得出,恐龙和翼龙一样和陆地生态系统一样在最初的几天内死于大火,海啸及风暴。
年代较晚的一批非鸟恐龙化石发现于K-Pg层下方13厘米处,是一批恐龙角化石,属于一种植食性恐龙,有可能是著名的三角龙的,因此它们可能是最后灭绝的恐龙。然而,成功穿越k-pg界线的恐龙依然存在。
较小的兽脚亚目恐龙可以躲避在天然洞穴(洞穴、大树干、水下/地下),藏在河流小溪边,以及非鸟类恐龙穴居的新证据使得这些动物中至少有一部分能够在一段时间内经受住灾难的影响。2003年,资深作者(JDS)和他的团队,由国家地理协会赞助探险时,首次发现了丹尼阶(古新世第一个阶)非鸟类恐龙的记录,在位于新西兰克赖斯特彻奇以东865公里的查塔姆岛上。较大的兽脚亚目恐龙遗骸包括兽脚亚目足指骨II-1(或III-1), 18.5 cm长,10.2厘米宽,可能属于一种中型的兽脚亚目,其他的骨头(脊椎,部分胫骨)代表这条恐龙长达4米。因此,有可能在西南太平洋的森林地带,有较小的兽脚亚目恐龙幸存了下来。
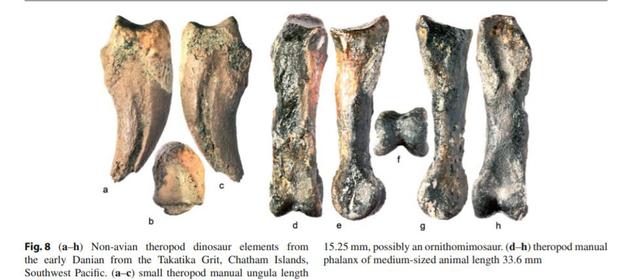 发现的兽脚类恐龙骨骼
发现的兽脚类恐龙骨骼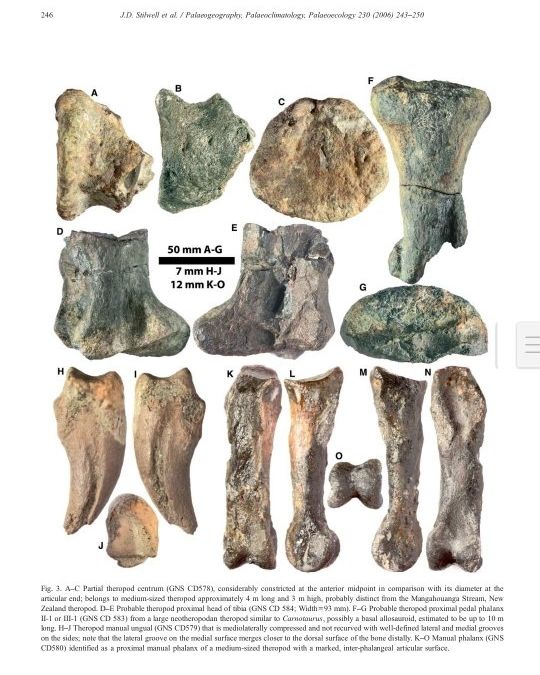 这批恐龙化石生存时间在80ma左右。
这批恐龙化石生存时间在80ma左右。中国科学家赵资奎在南雄地区白垩纪﹣古近纪灭绝事件(K/T)边界以外发现的恐龙蛋,说明陨石撞击理论并不适用于南雄地区的恐龙灭绝。根据他对恐龙蛋的研究,气候变化极大地影响了恐龙的食物链和繁殖过程,导致恐龙在20万到30万年的时间里逐渐灭绝,这也是非鸟类恐龙目前推测较长的延续时间。
中生代鸟类也是相似的情况。在K-Pg边界30万年以内的古代鸟类共鉴定出17种,包括反鸟,鱼鸟, 黄昏鸟,和一种类似神翼鸟的鸟类。这里描述的大多数鸟类都是今鸟类而不是反鸟。
这些末日前夕的鸟类是最近已知的最多样化的白垩纪鸟,包括较小的形式和一些中生代已知的最大的飞禽,虽然大小差距比现代要小。丰富的多样性表明鸟类在白垩纪末日之前有过一次大规模的辐射。但没有一种能确定地与新鸟类有关。其中一种,今鸟C(Ornithurine C),是已知的唯一的一种成功跨越K-Pg边界活到丹尼阶的鸟类。
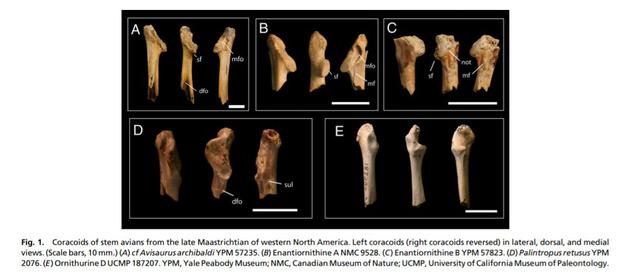 今鸟c(图中c)是已知的唯一的一种已跨越K-Pg边界的鸟类。
今鸟c(图中c)是已知的唯一的一种已跨越K-Pg边界的鸟类。另一个有较明确延续时间的是菊石。在马斯特里赫特地区的最后0.5ma,菊石仍旧较为繁盛,拥有中生代所有的四个亚目,包括六个超科、31(亚)属和57种。最近的数据表明,菊石一直存活在边界上。与恐龙同样的,它们也没有在灭绝后立刻消失。
Surlyk和Nielsen(1999)的一篇论文,《最后的菊石?》挑战了菊石灭绝于白垩纪末的理论,虽然他们没有详细的证据证明它们存活下来。他们的假设后来有了马查尔斯基(2002)的支持,他在丹尼阶发现了白垩纪末期有名的杆菊石和船菊石(均为异形菊石),尽管数量与马斯特里赫特时代晚期相比少得多。在他们之前,也有人发表过丹尼阶菊石的论文,但最后均被辨认为再沉积的结果,但这次,菊石周围没有其他白垩纪生物,而只有新生代特征的藻类,有孔虫和腕足类等,指示菊石也同样延续至丹尼阶,年约200kyr(1 kyr=1000年)。同样,以它们为食的沧龙和蛇颈龙也可能生存至此,但极有可能因为不耐饥饿死于最初的两年内。箭石也生活到古近纪,甚至延续到始新世。
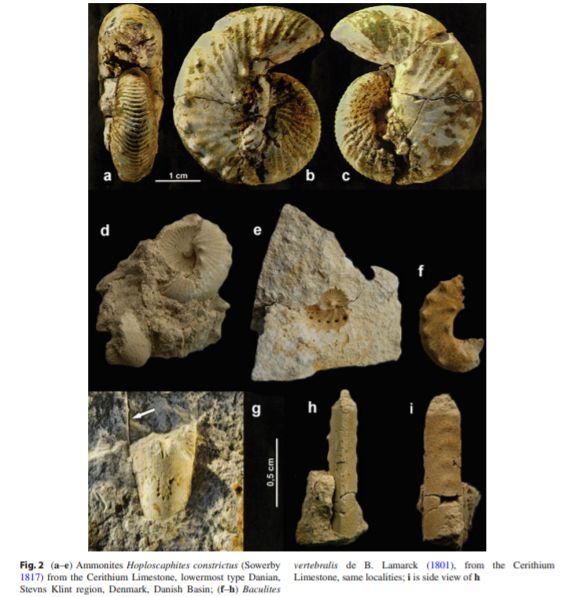 丹尼阶发现的船菊石
丹尼阶发现的船菊石 杆菊石
杆菊石起源于中三叠世的深海海百合Roveacrinids在白垩纪-古近纪存活了下来,尽管海百合似乎与海洋生物多样性的巨大变化没有关系。在白垩纪-古近纪(K-Pg)界线灭绝事件中,被认为是海百合类的代表的Roveacrinida目在这个时期灭绝了。但波兰的丹尼阶(古新世早期)保存完好的化石证明这些海百合存活到最早新生代。类似的还有喙头类的Opisthodontia,蚓蜥amphisbaenians,腹足类的Aporrhaidae,反羽蟹Costacopluma 以及苔藓虫。当然,这些类群没有一个活过丹尼阶。
中生代的生物就以它们的最终死去为止,画上了句号。
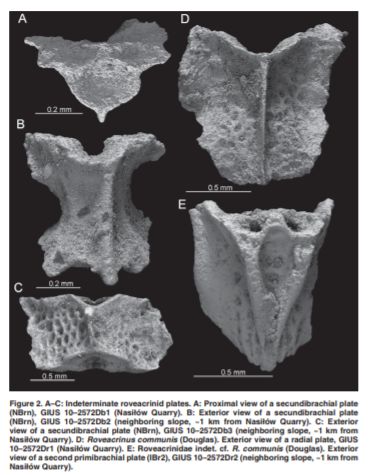 丹尼阶海百合
丹尼阶海百合 腹足类和喙头蜥
腹足类和喙头蜥 反羽蟹
反羽蟹参考文献
[1]Bralower T J, Paull C K, Mark Leckie R。 The Cretaceous-Tertiary boundary cocktail: Chicxulub impact triggers margin collapse and extensive sediment gravity flows[J]。 Geology, 1998, 26(4): 331-334。
[2]Morgan J, Warner M, Brittan J, et al。 Size and morphology of the Chicxulub impact crater[J]。 Nature, 1997, 390(6659): 472-476。
[3]Keller G, Adatte T, Stinnesbeck W, et al。 Chicxulub impact predates the KT boundary mass extinction[J]。 Proceedings of the National Academy of Sciences, 2004, 101(11): 3753-3758。
[4]Kring D A。 The Chicxulub impact event and its environmental consequences at the Cretaceous–Tertiary boundary[J]。 Palaeogeography, Palaeoclimatology, Palaeoecology, 2007, 255(1-2): 4-21。
[5]Schulte P, Alegret L, Arenillas I, et al。 The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary[J]。 Science, 2010, 327(5970): 1214-1218。
[6]GrajalesNishimura J M, CedilloPardo E, RosalesDomínguez C, et al。 Chicxulub impact: The origin of reservoir and seal facies in the southeastern Mexico oil fields[J]。 Geology, 2000, 28(4): 307-310。
[7]Vellekoop J, Sluijs A, Smit J, et al。 Rapid shortterm cooling following the Chicxulub impact at the CretaceousPaleogene boundary[J]。 Proceedings of the National Academy of Sciences, 2014, 111(21): 7537-7541。
[8]Sharpton V L, Burke K, Camargo-Zanoguera A, et al。 Chicxulub multiring impact basin: Size and other characteristics derived from gravity analysis[J]。 Science, 1993, 261(5128): 1565-1567。
[9]Robertson D S, Lewis W M, Sheehan P M, et al。 K‐Pg extinction: Reevaluation of the heat‐fire hypothesis[J]。 Journal of Geophysical Research: Biogeosciences, 2013, 118(1): 329-336。
[10]Grossnickle D M, Newham E。 Therian mammals experience an ecomorphological radiation during the Late Cretaceous and selective extinction at the KPg boundary[J]。 Proceedings of the Royal Society B: Biological Sciences, 2016, 283(1832): 20160256。
[11]Robertson D S, Lewis W M, Sheehan P M, et al。 KPg extinction patterns in marine and freshwater environments: the impact winter model[J]。 Journal of Geophysical Research: Biogeosciences, 2013, 118(3): 1006-1014。
[12]Apesteguía S, Gómez R O, Rougier G W。 The youngest South American rhynchocephalian, a survivor of the K/Pg extinction[J]。 Proceedings of the Royal Society B: Biological Sciences, 2014, 281(1792): 20140811。
[13]Salamon M A, Gorzelak P, Ferré B, et al。 Roveacrinids (Crinoidea, Echinodermata) survived the Cretaceous-Paleogene (K-Pg) extinction event[J]。 Geology, 2010, 38(10): 883-885。
[14]Longrich N R, Tokaryk T, Field D J。 Mass extinction of birds at the Cretaceous–Paleogene (KPg) boundary[J]。 Proceedings of the National Academy of Sciences, 2011, 108(37): 15253-15257。
[15]Longrich N R, Bhullar B A S, Gauthier J A。 Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary[J]。 Proceedings of the National Academy of Sciences, 2012, 109(52): 21396-21401
[16]Longrich N R, Bhullar B A S, Gauthier J A。 Mass extinction of lizards and snakes at the Cretaceous–
Paleogene boundary[J]。 Proceedings of the National Academy of Sciences, 2012, 109(52): 21396-21401
[17]Krug A Z, Jablonski D, Valentine J W。 Signature of the end-Cretaceous mass extinction in the modern biota[J]。 Science, 2009, 323(5915): 767-771。
[18]Hyžný M, Perrier V, Robin N, et al。 Costacopluma (Decapoda: Brachyura: Retroplumidae) from the Maastrichtian and Paleocene of Senegal: a survivor of K/Pg events[J]。 Cretaceous Research, 2016, 57: 142-156。
[19]Machalski M, Heinberg C。 Evidence for ammonite survival into the Danian (Paleogene) from the Cerithium Limestone at Stevns Klint, Denmark[J]。 Bulletin of the Geological Society of Denmark, 2005, 52(9)。
[20]Maor R, Dayan T, Ferguson-Gow H, et al。 Temporal niche expansion in mammals from a nocturnal ancestor after dinosaur extinction[J]。 Nature ecology & evolution, 2017, 1(12)。
[21]Vellekoop J, Smit J, van de Schootbrugge B, et al。 Palynological evidence for prolonged cooling along the Tunisian continental shelf following the K–Pg boundary impact[J]。 Palaeogeography, Palaeoclimatology, Palaeoecology, 2015, 426: 216-228。
[22]Dameron S N, Leckie R M, Clark K, et al。 Extinction, dissolution, and possible ocean acidification prior to the Cretaceous/Paleogene (K/Pg) boundary in the tropical Pacific[J]。 Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 485: 433-454。
[23]Sibert, E。 C。, & Norris, R。 D。 (2015)。 New Age of Fishes initiated by the Cretaceous−Paleogene mass extinction。 Proceedings of the National Academy of Sciences, 112(28), 8537–8542。 doi:10.1073/pnas.1504985112
[24]Jeffrey D。 Stilwell;Eckart H?kansson.Survival, but…! New Tales of ‘Dead Clade Walking’ from Austral and Boreal Post-K–T Assemblages[J]。Earth and Life.2012:795-810。



“掌”握科技鲜闻 (微信搜索techsina或扫描左侧二维码关注)











